
Biología:
ESTRUCTURA Y CARACTERÍSTICAS DE LOS VIRUS
Todo virus está formado por una envuelta proteica: la cápsida y por un ácido nucleico; además, algunos virus más complejos pueden tener una envoltura membranosa de lípidos y proteínas.
Los virus son muy pequeños y sólo son visibles mediante microscopía electrónica. Su tamaño oscila desde los 10 nm, en los pequeños virus de la poliomielitis, hasta los 300 nm en el virus de la viruela, el mosaico del tabaco -TMV- y otros. Se diferencian entre ellos, además de por el tamaño, por las características estructurales de la cubierta (la cápsida), por la naturaleza de su ácido nucleico, el modo de penetración en la célula hospedadora y el mecanismo de replicación.
3.1) Constitución y morfología de la cápsida
Todos los virus presentan, sin excepción, una envoltura proteica, denominada, cápsida, compuesta por el ensamblaje de una o varias subunidades proteicas llamadas capsómeros, dispuestas a menudo en varias capas concéntricas.
La geometría de la cápsida es uno de los criterios que permite clasificar los virus en cuatro grupos: icosaédricos, helicoidales, complejos y con envoltura.
* Icosaédricos: son los virus de aspecto esférico, cuya cápsida adopta la estructura de un icosaedro (poliedro de 20 caras triangulares, 30 aristas y 12 vértices); por ejemplo: los adenovirus, el virus de la polio y los picornavirus.
* Helicoidales o cilíndricos: están representados por el virus del mosaico del tabaco y el virus de la rabia; presentan un aspecto alargado, que en realidad corresponde a un cilindro hueco, donde los capsómeros se ensamblan siguiendo un ordenamiento helicoidal, similar a los peldaños de una escalera de caracol.
* Complejos, como bacteriófagos (virus parásitos de bacterias) que parecen adoptar las dos estructuras anteriores. Al igual que los icosaédricos poseen una región icosaédrica llamada cabeza donde se aloja el ADN y una cola formada por una banda de simetría helicoidal en cuyo interior se encuentra un eje tubular. La cola está terminada en un conjunto de fibras y espinas caudales que constituyen el sistema de anclaje del virus a la bacteria a la que infecta.
* Virus con envoltura membranosa: La mayoría de los virus animales, como los de la gripe, la viruela, la hepatitis, el virus del SIDA, etc. poseen, además de la cápsida, una envoltura membranosa que no es mas que un fragmento de la membrana plasmática de la célula hospedadora que el virus arrastra al abandonarla mediante un proceso de gemación. La bicapa lipídica que forma esta envoltura posee un conjunto de glucoproteínas codificadas por el virus y dispuestas hacia el exterior, a modo de espículas, que constituyen su sistema de anclaje en los receptores de membrana de las células hospedadoras y, por tanto, median en el mecanismo de penetración por endocitosis o por fusión de membranas. La envoltura membranosa es muy importante desde el punto de vista inmunológico
3.2) El ácido nucleico
Es el componente esencial del virus y puede ser ADN monocatenario, por ejemplo, en el fago O-X-174, o ADN bicatenario, como el fago T4 , y los adenovirus; pero también existen virus con ARN bicatenario (los reovirus) y otros portadores de ARN monocatenario, como es el caso de los virulentos retrovirus, entre los que se encuentran el de la gripe, el sarampión, la rabia, el SIDA y determinados virus oncógenos causantes de ciertos tipos de cáncer (sarcoma de Rous, determinadas leucemias, etc.). Este último grupo contiene, además de los otros componentes mencionados, un enzima particular llamado retrotranscriptasa o transcriptasa inversa, que le va a permitir transcribir su ARN en un ADN dentro de la célula infectada.
MECANISMOS DE REPLICACIÓN: CICLO VITAL DE LOS VIRUS
Aunque el genoma de un virus contiene escaso número de genes, es suficiente para inhibir la expresión génica de la célula hospedadora y obligarla a transcribir y traducir su breve mensaje. El modo de penetración , los mecanismos y los compartimentos celulares utilizados para la replicación, son diferentes en los distintos tipos de virus. De todos ellos, se pondrán como ejemplo el de los retrovirus y los bacteriófagos.
a) Ciclo vital de un retrovirus: El VIH causante del SIDA.
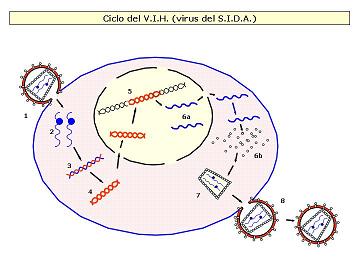
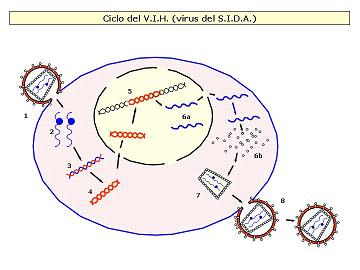
Los retrovirus son un grupo muy especial de virus animales. Son virus cuyo ácido nucleico es ARN, poseen envoltura y el enzima transcriptasa inversa.
EL VIH es un retrovirus relativamente complejo. Está constituido por una membrana lipídica con glucoproteínas dispuestas al exterior a modo de espículas. En el interior encontramos una cápsida proteica que encierra el material genético, formado por dos moléculas de ARN monocatenario y se encuentran ligadas, cada una de ellas, a una molécula de una enzima, la transcriptasa inversa.
Información: El VIH ataca preferentemente a los linfocitos T4. Las fases de este proceso son:
1ª) Contacto entre las espículas de su envoltura membranosa y los receptores de la célula hospedadora. Estas permiten la fusión de membranas, introduciendo en su interior la cápside con el material genético.
2ª) Una vez en el interior, el virus se despoja de su cápsida protéica y quedan libres las hebras de ARN y la enzima retrotranscriptasa que transporta.
3ª) La retrotranscriptasa, también llamada transcriptasa inversa, primero hace una copia en ADN de la cadena de ARN, es decir, invierte el proceso normal de transcripción de ADN a ARN, originando una hélice híbrida ARN-ADN.
4ª) La hélice híbrida ARN-ADN es utilizada por la misma enzima para generar una doble hélice de ADN (previa degradación del ARN).
5ª) Las dobles cadenas de ADN víricas entran en el núcleo y se insertan en el cromosoma celular, donde puede permanecer en estado latente en forma de provirus durante un tiempo más o menos prolongado.
6ª) Finalmente se transcriben y se traducen utilizando la maquinaria metabólica de la célula y origina nuevas copias de ARN vírico, proteínas de la cápsida y de la envoltura y enzimas retrotranscriptasas.
7ª) Estos componentes se ensamblan, y...
8ª) los virus abandonan la célula mediante un proceso de gemación que les permite adquirir de nuevo su recubrimiento membranoso.
Todos estos procesos pueden ser lentos, originando tan sólo un descenso de la actividad metabólica del hospedador, o rápidos, con lo que la salida masiva de virus termina con la lisis de la célula.
b) Ciclo vital del fago T4.
El bacteriófago T4 es un virus complejo con una cabeza icosaédrica y una cola en la que hay una placa basal y fibras de fijación. El genoma se compone de una molécula de ADN bicatenaria que se encuentra profusamente empaquetada dentro de la cabeza.
El fago se fija en la pared bacteriana, en las regiones denominadas puntos de adherencia, a través de los cuales inyecta su ADN mediante la contracción de la vaina de la cola. Una vez en el protoplasma bacteriano, el ADN puede seguir dos caminos: multiplicarse y originar nuevos virus (vía lítica), con lo que se produce la destrucción de la bacteria, o integrarse en el cromosoma bacteriano y adoptar la forma de profago (vía lisogénica).
i) Ciclo lítico.
1) Fijación y entrada: Inicialmente el bacteriófago fija su cola a receptores específicos de la pared de la bacteria donde una enzima, localizada en la cola del virus, debilita los enlaces de las moléculas de la pared. A continuación el fago contrae la vaina helicoidal, lo que provoca la inyección del contenido de la cabeza a través del eje tubular de la cola del fago: el ácido nucleico penetra en la célula.
2) Multiplicación: Una vez dentro el ADN del virus, utilizando nucleótidos y la enzima ARNpolimerasa de la bacteria, dirige la síntesis de gran cantidad de ARNm viral. Este ARNm viral sirve de base para la síntesis de proteínas del virus (capsómeros, endonucleasas, endolisinas). El ADN vírico, utilizando los complejos enzimáticos de la bacteria, se replica muchas veces. Tanto los ácidos nucleicos replicados, como el resto de los componentes víricos que se han sintetizado, se ensamblan dando lugar a nuevos virus.
3) Lisis y liberación. En una bacteria pueden formarse unos 100 bacteriófagos que salen al exterior debido a la acción de la endolisina, enzima que lisa la pared bacteriana. Debido a ello se produce la ruptura de la pared bacteriana y la muerte de la célula. Los virus quedan libres para infectar nuevas células.
ii) Ciclo lisogénico.
No siempre se produce la lisis inmediata de la célula. Hay fagos atemperados o atenuados que se integran en el ADN bacteriano por entrecruzamiento de dos regiones idénticas del fago y de la bacteria, del mismo modo a como ocurre en los plásmidos. Estos fagos integrados se denominan profagos, y se replican pasivamente con el ADN de la bacteria. Las bacterias capaces de establecer esa relación con los fagos atenuados se denominan lisogénicas.
El ADN del profago puede permanecer en forma latente durante varias generaciones de la bacteria, hasta que un estímulo induzca la separación del profago que iniciará un ciclo lítico típico. Mientras la célula posea el ADN profago será inmune frente a infecciones de este mismo virus. Otros virus que no son bacteriófagos pueden también tener ciclos lisogénicos.
CLASIFICACIÓN DE LOS VIRUS
MECANISMOS DE TRANSCRIPCIÓN DE LOS VIRUS EN FUNCIÓN DE SU MATERIAL GENÉTICO.
VIROIDES
Son extremadamente sencillos y forman un escalón inferior a los virus. Son simplemente genomas desnudos, ARN de una cadena (pero en forma de horquilla, pues hay complementariedad entre sus bases, simulando un ARN doble para protegerse de los enzimas hidrolíticos celulares que atacan a los ARN simples) y no presentan cápsida proteica. Solamente causan enfermedades en los vegetales. Los viroides han producido pérdidas económicas importantes en cultivos de patata en USA y en cocoteros en Filipinas.
Son de menor tamaño que cualquiera de los genomas víricos conocidos, pero suficiente para poder codificar una proteína, pero no se cree que lo hagan ya que el ARN de los viroides carece de señales que se necesitan para la traducción del ARN a una proteína. Por lo tanto su información no se traduce, solo se replica. Parece probable que sea la ARNpolimerasa del hospedador, que está en el núcleo de las plantas, la que replica el genoma del viroide. No está claro cómo se transmiten entre células, dada la dificultad que opone la pared celular de las células vegetales a las que infectan.
LOS PRIONES: De estos "organismos" sabemos aún menos. Se descubren en 1983 como agentes causantes de afecciones neuronales esporádicas. Ahora aumenta su interés debido al mal de las vacas locas.
Es una partícula infecciosa proteínica (proteína patológica). Las pruebas obtenidas hasta el momento parecen indicar que el prión carece de ácido nucléico.
Se conocen dos enfermedades causadas por priones: La Tembladera, una alteración neurológica de ovejas y cabras, conocida desde el siglo XVII y la enfermedad de Creutzfeld-Jacob, una rara demencia humana. Los priones también se consideran agentes probables de otras enfermedades humanas que afectan al sistema nervioso: el Kuru, observado sólo en tribus de Nueva Guinea, asociándose al canibalismo tradicional (la enfermedad fue desapareciendo conforme cesaban las prácticas necrófagas).
La enfermedad de Creutzfeld-Jacob en individuos menores de 35 años se relacionó con el consumo de subproductos de vacas enfermas, que estaban alimentadas con piensos fabricados con restos de ovejas con tembladera.
La infección por priones no provoca una respuesta inmunitaria, debido a que el prión está dentro de nuestras propias células. El agente causante es una proteína propia de la membrana plasmática de las neuronas. Se sabe que está codificada por un gen del cromosoma 20. Esta proteína sufre una alteración que la convierte en patológica (prión) Las proteínas defectuosas actúan como agentes infecciosos que cambian las proteínas normales en defectuosas. La aparición de la demencia es consecuencia de que se acumulan cristalizadas en las neuronas provocando su destrucción y muerte.
Comparando las dos proteínas, normal y patológica, se comprueba que tienen la misma secuencia de aminoácidos (estructura primaria), pero tienen un plegamiento distinto.
Se han encontrado casos de transmisión hereditaria de la enfermedad, debido a una mutación puntual que implica modificación en la estructura primaria de la proteína, sustituyéndose una prolina por una leucinaFuente